 W
WAsilisaurus – rodzaj dinozauromorfa z grupy Silesauridae żyjącego w środkowym triasie na obecnych terenach Afryki. Został opisany w 2010 roku przez Sterlinga Nesbitta i współpracowników w oparciu o przednią część kości zębowej wraz z zębami odkrytą w datowanych na anizyk osadach ogniwa Lifua w złożach Manda w pobliżu tanzańskiego miasta Litumba Ndyosi. Odnaleziono jednak izolowane szczątki należące do co najmniej czternastu osobników, składające się w sumie na niemal kompletny szkielet – jedynie niektóre części czaszki oraz nadgarstki pozostają nieznane. Według Nesbitta i współpracowników (2010) większość skamieniałości należała do osobników młodocianych, które osiągnęły 60–80% maksymalnych rozmiarów; na tej podstawie, oraz na podstawie porównań ze szkieletem silezaura, autorzy szacowali, że dorosli przedstawiciele gatunku A. kongwe osiągali około 2–3 m długości ciała. Czaszka ma podobne proporcje jak czaszka silezaura i podobnie jak u niego oraz u Sacisaurus kość zębowa jest ostro zakończona. Szacowana długość kości ramiennej i promieniowej sugeruje, że Asilisaurus miał wydłużone kończyny przednie, jak Silesaurus. Panewka stawu biodrowego była zamknięta, jak u wszystkich przedstawicieli Ornithodira nienależących do dinozaurów. Kość skokowa i kość piętowa to u Asilisaurus odrębne struktury, w przeciwieństwie do pterozaurów, teropodów i Lagerpetidae, u których były one na wpół skostniałe.
 W
WAwipes – żyjący w środkowym triasie (ladyn) archozaur o niepewnej pozycji systematycznej. Jego nazwa rodzajowa oznacza "ptasia stopa"; nawiązuje ona do podobieństwa kości śródstopia tego zwierzęcia do ptasiej. Znany wyłącznie z trzech bardzo słabo zachowanych fragmentów kości śródstopia mierzących ok. 80 mm, odkrytych niedaleko Dillstedt w Turyngii (Niemcy). Początkowo klasyfikowany jako ceratozaur lub celurozaur; Rauhut i Hungerbühler (2000) uznali Avipes dillstedtianus za nomen dubium i stwierdzili, że z dużą pewnością można jedynie zaliczyć go do archozaurów, natomiast bliższa klasyfikacja jest niemożliwa.
 W
WAzendozaur (Azendohsaurus) – rodzaj roślinożernego archozauromorfa żyjącego w późnym triasie na obecnych terenach Afryki. Gatunek typowy rodzaju, Azendohsaurus laaroussi, został opisany na podstawie kości zębowej zawierającej zęby oraz izolowanych zębów. Później przypisano do niego także inne znalezione w Maroku szczątki, złożone z kręgów piersiowych, kości obręczy barkowej oraz przednich i tylnych kończyn. Początkowo na podstawie podobieństw do zębów fabrozaura i likorina uznany za roślinożernego dinozaura ptasiomiednicznego, jednak późniejsi autorzy sugerowali, że jest on prozauropodem. Nowsze badania wskazują jednak, że Azendohsaurus nie należy do dinozaurów ani w ogóle archozaurów, lecz jest bazalnym archozauromorfem, być może blisko spokrewnionym z trilofozaurami. Stosunkowo kompletne szczątki pochodzące z przełomu środkowego i późnego triasu Madagaskaru, opisane w 2010 roku jako Azendohsaurus madagaskarensis, potwierdzają status azendozaura jako bazalnego archozauromorfa, u którego występowały niektóre cechy uznawane wcześniej za diagnostyczne dla zauropodomorfów. Szczątki A. laaroussi pochodzą z datowanych na karnik osadów formacji Argana w górach Atlas, w pobliżu wioski Azendoh w regionie Marrakesz-Tansift-Al-Hauz. Z tej samej formacji pochodzą odciski stóp przypisywane ptasiomiednicznym i teropodom. Azendozaur osiągał prawdopodobnie około 1,8 m długości.
 W
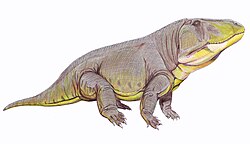
WBatrachotomus – rodzaj prehistorycznego archozaura. Skamieniałości znaleziono w południowych Niemczech, datuje się je na środkowotriasowy ladyn, około 242-235 milionów lat temu. Rodzaj opisał paleontolog David J. Gower 22 lata po odkryciu.
 W
WBrachyrhinodon – rodzaj lepidozaura z rodziny hatteriowatych (Sphenodontidae). Jego skamieniałości odkryto w datowanych na karnik osadach formacji Lossiemouth Sandstone w północnej Szkocji. Jest to jeden z najstarszych znanych sfenodontów. Brachyrhinodon osiągał 15–20 cm długości. Miał ostre zęby służące prawdopodobnie do ścinania niewielkich roślin. Jest blisko spokrewniony z rodzajami Polysphenodon i Clevosaurus, wraz z którymi należy do kladu Clevosauridae. Zarówno Polysphenodon, jak i Brachyrhinodon miały tępe pyski, jednak jak wykazali Fraser i Benton (1989) stopień skrócenia pyska u obu tych form był przeszacowany ze względu na zniekształcenia znanych skamieniałości. Wydłużona okolica skroniowa oraz długość regionu międzyoczodołowego większa niż długość płyty ciemieniowej są cechami zaawansowanymi wśród hatteriowatych. Polysphenodon w obu przypadkach wykazuje cechy plezjomorficzne, zaś Brachyrhinodon – zaawansowane.
 W
WClevosaurus – rodzaj lepidozaura z rodziny hatteriowatych (Sphenodontidae), z grupy Clevosauridae.
 W
WDinocefalozaur (Dinocephalosaurus) – rodzaj długoszyjego wodnego gada z rzędu Prolacertiformes, żyjącego w triasie, około 244 mln lat temu. Jego nazwa oznacza „jaszczur o straszliwej głowie” – ze względu na fakt, że pierwszą odnalezioną skamieniałością była czaszka odkryta w 2002 w pobliżu Xinmin w chińskiej prowincji Guizhou. Szczątki drugiego osobnika zostały odkryte niedaleko miejsca znalezienia pierwszego i zawierały głowę i większość szkieletu pozaczaszkowego oprócz ogona. Dinocephalosaurus jest pierwszym protorozaurem odkrytym w środkowotriasowych osadach w Chinach.
 W
WDiodorus – rodzaj archozaura z grupy Dinosauriformes żyjącego w późnym triasie na obecnych terenach Maroka.
 W
WDrepanosauridae – rodzina zauropsydów z grupy diapsydów; żyły w okresie późnego triasu na terenach dzisiejszej Europy, Ameryki Północnej i Azji (Kyrgyzsaurus). Zdaniem większości naukowców zwierzęta z tej rodziny prowadziły nadrzewny tryb życia, prawdopodobnie zajmując nisze ekologiczne dzisiejszych kameleonów; rodzaj Hypuronector, zdaniem autorów jego opisu, mógł jednak prowadzić ziemnowodny tryb życia. Posiadały przeciwstawne palce obydwu par kończyn, a niektóre miały "pazur" na końcu ogona przydatny przy wspinaczce. Wymarły pod koniec triasu.
 W
WDrepanosaurus jest nazwą rodzajową nadrzewnego diapsyda żyjącego w późnym triasie na terenach obecnej Europy i Ameryki Północnej. Był to pierwszy opisany gad swojej rodziny, która w związku z tym nazwana została Drepanosauridae. Megalankozaur żył w tym samym czasie i na tym samym terenie co drepanozaur, ale był od niego dużo mniejszy.
 W
WDromomeron – rodzaj archozaura z kladu Dinosauromorpha, obejmującego dinozaury i blisko spokrewnione z nimi prymitywniejsze archozaury. Dromomeron był jednym z najprymitywniejszych przedstawicieli tego kladu; analizy kladystyczne sugerują, że jego najbliższymi znanymi krewnymi są starsze o kilkanaście milionów lat Lagerpeton z Argentyny oraz Ixalerpeton z Brazylii. Gatunkiem typowym rodzaju jest nazwany w 2007 Dromomeron romeri, nazwany na cześć paleontologa Alfreda Romera. W 2009 opisano drugi gatunek należący do rodzaju Dromomeron – D. gregorii, a w roku 2016 trzeci gatunek – D. gigas. Szacuje się, że gatunek typowy osiągał długość około 1 metra.
 W
WErythrosuchus jest nazwą drapieżnego archozaura, żyjącego we wczesnym i środkowym triasie na terenie obecnej południowej Afryki, która wchodziła wtedy w skład superkontynentu Pangei.
 W
WEuperkeria – rodzaj niewielkiego archozauromorfa z grupy Archosauriformes. Żył we wczesnym triasie, około 248 milionów lat temu. Jego szczątki odkryto tylko na jednym stanowisku w regionie Karoo. Znane są skamieniałości zaledwie kilku osobników. Nazwa Euparkeria nawiązuje do nazwiska W. K. Parkera.
WEuparkeriidae – rodzina niewielkich drapieżnych bazalnych archozauromorfów z kladu Archosauriformes. Jej przedstawiciele żyli od wczesnego do środkowego triasu (anizyk). Ich skamieniałości zostały odnalezione w południowej Afryce, Polsce i Rosji. W przeciwieństwie do siostrzanej rodziny Erythrosuchidae przedstawiciele Euparkeriidae byli niewielcy, smukli i prawdopodobnie poruszali się na dwóch nogach.
 W
WGarjainia – rodzaj drapieżnego archozauromorfa z wczesnego triasu (olenek), obejmujący gatunki opisane na podstawie skamieniałości odkrytych na obszarze Rosji i Republiki Południowej Afryki. Długość jej ciała wynosiła 1,5-1,8 metra. Zwierzę uważane jest za blisko spokrewnione z południowoafrykańskim erytrozuchem.
 W
WHyperodapedon – rodzaj archozauromorfa z grupy rynchozaurów (Rhynchosauria).
 W
WHypsognathus jest nazwą rodzajową roślinożernego gada z grupy anapsydów, żyjącego pod koniec triasu. Jego skamieniałości znaleziono na terenie stanu New Jersey i Nowej Szkocji. W związku z tym uważa się, że zwierzę to było rozpowszechnione na obszarze środkowo-zachodniej Pangei.
 W
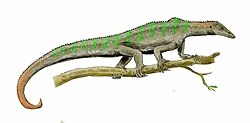
WIkarozaur (Icarosaurus) – rodzaj diapsyda z rodziny Kuehneosauridae. Żył w późnym triasie na terenie dzisiejszej Ameryki Północnej. Jego szczątki odkryto w formacji Lockatong w stanie New Jersey. Został opisany w 1966 roku przez Edwina Harrisa Colberta na podstawie niekompletnego szkieletu niezawierającego części ogona, kilku żeber i fragmentów kończyn. Ikarozaur był niewielkim zwierzęciem, osiągającym długość około 10 cm od głowy do nasady ogona. Miał puste w środku kości oraz duże oczodoły. Podobnie jak jego bliscy krewni Kuehneosaurus i Kuehneosuchus, ikarozaur był zdolny do szybowania na krótkie dystanse przy użyciu „skrzydeł” zbudowanych z wydłużonych, pokrytych skórą żeber, z wypukłą górną, a wklęsłą dolną powierzchnią, tworzących profil lotniczy odpowiedni do szybowania. Na podstawie stosunku obciążenia jednostkowego powierzchni „skrzydła” do zdolności szybowniczych u współczesnych jaszczurek z rodzaju Draco McGuire i Dudley (2011) ocenili, że Icarosaurus jest prawdopodobnie najlepiej przystosowany do szybowania spośród wszystkich znanych kręgowców nielatających aktywnie.
 W
WIxalerpeton – rodzaj wczesnego dinozauromorfa należący do kladu Lagerpetidae. Żył w późnym triasie, ok. 230 mln lat temu, na terenach dzisiejszej Ameryki Południowej. Był stosunkowo niewielkim zwierzęciem, mierzącym ok. 1 m długości. Ixalerpeton wykazywał mozaikę cech pierwotnych dla archozaurów, jak występowanie dwóch kręgów krzyżowych czy zamknięta panewka stawu biodrowego, oraz zaawansowanych, jak obecność przedniego zagłębienia bębenkowego mózgoczaszki, które wśród dinozauromorfów występuje u przedstawicieli bardziej zaawansowanego kladu Dinosauriformes.
 W
WKuehneosauridae – rodzina diapsydowych szybujących gadów żyjących w triasie. Charakteryzowały się rozpiętymi na bok, długimi żebrami, które były dla nich główną powierzchnię nośną. Innymi wspólnymi cechami były połączone w jedno nozdrza i zredukowane kości łuskowe.
WKühneozaur (Kuehneosaurus) – rodzaj wymarłych późnotriasowych gadów z Wielkiej Brytanii. Długi na 72 cm, miał „skrzydła” utworzone z żeber wystających na 14,3 cm, połączonych błoną, które pozwalały mu spowolnić spadek w czasie skoków po drzewach.
 W
WLagerpeton – rodzaj dinozauromorfa żyjącego w późnym triasie na terenach współczesnej Ameryki Południowej. Został opisany w 1971 roku przez amerykańskiego paleontologa Alfreda Romera w oparciu o niemal kompletną, połączoną stawowo kończynę tylną odkrytą w formacji Chañares w Argentynie. Rok później Romer przypisał do rodzaju Lagerpeton dwie kości udowe, strzałkowe, łokciowe, kość piszczelową i liczne kręgi, jednak było to kwestionowane przez późniejszych autorów – Sereno i Arcucci stwierdzili, że zasadne jest przypisanie jedynie dwóch kości udowych, charakteryzujących się hakowatą głową, wydłużonym czwartym krętarzem i powiększonym kłykciem strzałkowym, które są cechami typowymi dla lagerpetona. Lagerpeton miał długie i smukłe kończyny tylne – palce I i V były zredukowane, II znacznie krótszy od III, a III nieco krótszy od IV. Na podstawie długości kości udowej długość lagerpetona szacuje się na około 70 cm. Archozaur ten był dwunożnym zwierzęciem, prawdopodobnie poruszającym się za pomocą skoków.
 W
WLagozuch (Lagosuchus) – rodzaj niewielkiego archozaura z kladu Ornithodira, jedyny przedstawiciel rodziny Lagosuchidae. Żył w środkowym triasie na terenie dzisiejszej Argentyny. Nazwa Lagosuchus oznacza „króliczy krokodyl”.
 W
WLeptopleuron – rodzaj niewielkiego zauropsyda należącego do grupy Procolophonidae. Żył w późnym triasie na terenie współczesnej Europy. Jego skamieniałości odkryto w datowanych na karnik lub noryk osadach formacji Lossiemouth Sandstone w północnej Szkocji.
 W
WLewisuchus – rodzaj dinozauromorfa żyjącego w późnym triasie na obecnych terenach Ameryki Południowej. Być może należy do grupy Silesauridae.
 W
WLongiskwama (Longisquama) – rodzaj wymarłego diapsyda żyjącego w późnym triasie na obecnych terenach Azji. Został opisany w 1970 roku przez Aleksandra Szarowa w oparciu o niekompletny szkielet odkryty w Kirgistanie. Szarow przypisał go do grupy Pseudosuchia, należącej do archozaurów. Swoją teorię oparł na obecności u longiskwamy okien przedoczodołowego oraz żuchwowego, będących synapomorfiami archozaurów. Przynależność longiskwamy do Archosauriformes – mimo iż wspierana przez niektórych autorów – nie jest jednak pewna, gdyż obecności okna przedoczodołowego nie można potwierdzić, a domniemane okno żuchwowe jest wynikiem uszkodzenia skamieniałości – i tak zresztą znajduje się ono w innym miejscu niż u archozaurów. Ponadto longiskwamy nie łączą z archozaurami żadne inne cechy diagnostyczne. Uzębienie akrodontyczne oraz skostniałe kości międzyobojczykowe, występujące u longiskwamy, są z kolei cechami typowymi dla lepidozaurów. Najbardziej charakterystyczną cechą longiskwamy są długie twory naskórka znajdujące się na grzbiecie zwierzęcia. W 1987 roku Haubold i Buffetaut zasugerowali, że po ich rozłożeniu na boki mogły umożliwiać longiskwamie krótki lot szybowcowy, jednak teoria ta nie jest obecnie uważana za prawdopodobną. W 2000 roku Terry D. Jones i współpracownicy stwierdzili, że twory te są bardzo podobne do piór współczesnych ptaków i prawdopodobnie z nimi homologiczne, ocenili jednak, że stopień pokrewieństwa pomiędzy longiskwamą a ptakami pozostaje nieznany. David Unwin i Michael J. Benton (2001) oraz Robert Reisz i Hans-Dieter Sues (2000) stwierdzili jednak, że struktury interpretowane przez Jonesa i współpracowników jako „nieptasie pióra” są w rzeczywistości w wielu aspektach całkowicie odmienne od piór ptaków. Unwin i Benton odrzucili hipotezę o przynależności longiskwamy do archozaurów, a wraz z Alifanowem stwierdzili, że teorię mówiącą, iż Longisquama jest przodkiem ptaków należy odrzucić ze względu na brak dowodów na jej poparcie. Analiza filogenetyczna przeprowadzona w 2004 roku przez Phila Sentera sugeruje, że Longisquama nie tylko nie należy do archozaurów, lecz jest bazalnym przedstawicielem diapsydów, przypuszczalnie należącym do grupy Avicephala. Badania Sebastiana Voigta i współpracowników z 2009 roku nie wspierają kategoryzowania wydłużonych tworów na grzbiecie longiskwamy ani jako piór, ani jako łusek. Voigt i in. sugerują, że struktury te wykształciły się w dwustopniowym procesie przypominającym proces powstawania pióra i są przykładem niezwykłej plastyczności tworów naskórka występujących u zauropsydów.
 W
WMalerizaur (Malerisaurus) – rodzaj niewielkiego gada należącego do archozauromorfów. Żył w późnym triasie (karnik) na terenie dzisiejszych Indii oraz Ameryki Północnej. Pierwotnie zaliczony do grupy Prolacertiformes i rodziny Protorosauridae; z badań Nesbitta i współpracowników (2017) wynika jego przynależność do grupy Allokotosauria i do rodziny Azendohsauridae.
 W
WMarazuch (Marasuchus) – rodzaj niewielkiego triasowego archozaura z kladu Dinosauriformes, blisko spokrewnionego z dinozaurami. Nazwa Marasuchus wywodzi się od słów: mara – nazwy gryzoni żyjących w Ameryce Południowej – i od greckiego słowa souchos, oznaczającego krokodyla. Marasuchus był małym, smukłym, długoogoniastym archozaurem. Miał długi tułów i esowato wygiętą szyję i mierzył około 40 cm długości. Występował w późnym triasie, około 236–234 miliony lat temu, na obszarze dzisiejszej Argentyny.
WMegalancosaurus jest nazwą rodzajową diapsyda, żyjącego w górnym triasie na terenie obecnych północnych Włoch.
 W
WMiodentosaurus - rodzaj zauropsydów, do którego zaliczany jest:Miodentosaurus brevis Cheng et al., 2007
 W
WOrnithosuchus - drapieżny wymarły archozaur z grupy Crurotarsi. Żył w triasie około 220 mln lat temu na terenie dzisiejszej Szkocji. Długość ciała ok. 4 metrów, waga 70 kg.
 W
WOsmolskina – rodzaj bazalnego przedstawiciela Archosauriformes żyjącego we wczesnym triasie na terenie dzisiejszej Europy. Holotyp Osmolskina czatkowicensis, jedynego gatunku rodzaju, został odkryty w datowanych analizą biostratygraficzną na najprawdopodobniej wczesny olenek osadach w Czatkowicach niedaleko Krakowa. Obejmuje przednią część kości szczękowej z zachowanym wyrostkiem nosowym. Odkryto także dwieście kości czaszki, w tym kości puszki mózgowej oraz żuchwy, trzydzieści kręgów ze wszystkich odcinków kręgosłupa, pięć kompletnych kości biodrowych, trzydzieści kości kończyn oraz kilkaset kości szkieletu pozaczaszkowego i zębów. Długość czaszki osmolskiny wynosiła około 60 mm, jednak niektóre z odkrytych skamieniałości sugerują, że u większych osobników dochodziła ona do około 120 mm.
 W
WOzimek – rodzaj archozauromorfa żyjącego w późnym triasie na terenach dzisiejszej Europy.
 W
WPisanozaur (Pisanosaurus) – rodzaj archozaura z grupy Dinosauriformes żyjącego w okresie triasu na terenach dzisiejszej Ameryki Południowej. Jego szczątki znaleziono w Argentynie.
 W
WProlacerta – rodzaj wczesnego archozauromorfa przypominającego sylwetką i rozmiarami współczesne jaszczurki. Odnaleziona we wczesnotriasowych osadach Afryki i Antarktydy. Dawniej uznawana za ogniwo pośrednie między prymitywnymi permskimi diapsydami, takimi jak Youngina, a jaszczurkami. Poparte to było brakiem łuku jarzmowego, ale późniejsze badania wykazały, że absencja tej struktury jest cechą obecną już u wspólnego przodka lepidozaurów i archozaurów. Sugerowany kinetyzm czaszki również został zanegowany. Prolacerta została później przeniesiona do grupy Archosauromorpha, do nowo utworzonego rzędu Prolacertiformes, który okazał się polifiletyczny. Obecnie umiejscawia się ją blisko kladu Archosauriformes obejmującego m.in. archozaury. Znana z licznego materiału, obejmujących zarówno pozostałości czaszki jak i szkieletu pozaczaszkowego. Jej młodszym synonimem jest Pricea longiceps Broom i Robinson 1948 znana również z zony Lystrosaurus. Również Kadimakara Bartholomai, 1979 z wczesnego triasu Australii może okazać się synonimem prolacerty.
 W
WProlacertiformes – rząd diapsydów żyjących w permie i triasie. Grupę tę wyróżnił w 1945 roku Charles Camp, zaliczając do niej rodzaje Prolacerta, Protorosaurus, Tangasaurus i Youngina. W przeszłości Prolacertiformes/Protorosauria zaliczano do lepidozaurów, bywały nawet zaliczane do gadów łuskonośnych i uznawane za możliwych przodków współczesnych jaszczurek. Rodzaje Protorosaurus i Tanystropheus bywały też zaliczane do grupy Euryapsida i uznawane za bliskich krewnych zauropterygów. Analizy kladystyczne wykazały jednak, że Prolacertiformes w rzeczywistości należały do archozauromorfów.
 W
WProterozuch (Proterosuchus) – drapieżny gad z grupy Archosauromorpha, żyjący około 250 milionów lat temu, we wczesnym triasie. Być może występował już pod koniec permu. Należał do rodziny Proterosuchidae, obejmującej takie rodzaje jak chasmatozuch, kalizuch, czy tasmaniozaur.
 W
WRevueltosaurus – rodzaj archozaura z grupy Crurotarsi żyjącego w późnym triasie na terenie dzisiejszej Ameryki Północnej. Gatunek typowy rodzaju, Revueltosaurus callenderi, został opisany przez Adriana Hunta w 1989 roku w oparciu o niemal kompletny ząb z kości przedszczękowej. Hunt uznał R. callenderi za dinozaura ptasiomiednicznego. Bardziej kompletne szczątki, odnalezione w datowanych na noryk, około 209 mln lat, osadach formacji Chinle na terenie Parku Narodowego Skamieniałego Lasu i opisane w 2005 roku przez Williama Parkera i współpracowników, dowiodły, że Revueltosaurus w rzeczywistości nie był dinozaurem, lecz przedstawicielem kladu Crurotarsi, obejmującego archozaury bliżej spokrewnione z krokodylami niż z ptakami. Czaszka Revueltosaurus była długa i niska. W kości przedszczękowej znajdowało się co najmniej pięć zębów, zaś w szczękowej – dziewięć lub dziesięć zębodołów. Budowa stawu skokowego górnego jest typowa dla krokodyli. Kość udowa bardzo przypomina tę turfanozucha – ma duży czwarty krętarz i słabo wykształconą głowę. Cechy uzębienia, wykorzystane wcześniej do zaklasyfikowania Revueltosaurus do Ornithischia, są obecnie uznawane za konwergentne u dinozaurów ptasiomiednicznych i niektórych Crurotarsi. W późniejszych pracach również włączano Revueltosaurus do Crurotarsi, a nie Dinosauria. Niektóre analizy kladystyczne sugerują jego bliskie pokrewieństwo z Ornithosuchidae, a inne z aetozaurami.
 W
WRhynchosaurus – rodzaj archozauromorfa należący do grupy rynchozaurów. Jego przedstawiciele żyli w środkowym triasie na terenie dzisiejszej Europy. Obejmuje dwa gatunki: Rhynchosaurus articeps, będący gatunkiem typowym rodzaju, oraz Rhynchosaurus brodiei. Michael J. Benton opisał także trzeci gatunek, R. spenceri, jednak jest on obecnie uznawany za należący do odrębnego rodzaju Fodonyx.
 W
WSacisaurus – rodzaj archozaura z grupy Dinosauriformes żyjącego w późnym triasie na obecnych terenach Ameryki Południowej. Został opisany w oparciu o niekompletną lewą kość żuchwy, zawierającą większość kości zębowej, trzy zęby wykazujące przystosowania do roślinożerności oraz bezzębną część rostralną, mogącą stanowić odrębną parę skostnień. W datowanych na późny karnik lub wczesny noryk osadach formacji Caturrita w Agudo, w południowej Brazylii, odkryto również kilka kości czaszki, kręgów, łopatkę, kość miedniczna, kości kończyn tylnych oraz liczne zęby. Nazwa rodzajowa Sacisaurus pochodzi od imienia Saci – jednonogiej istoty z brazylijskiej mitologii – oraz greckiego słowa sauros, oznaczającego „jaszczur”, i odnosi się do faktu, iż odnaleziono tylko jedną kość udową zwierzęcia. Epitet gatunkowy agudoensis pochodzi od miejsca odkrycia szczątków.
 W
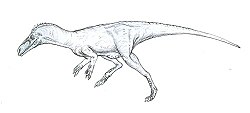
WSaltopus – rodzaj niewielkiego dwunożnego archozaura należącego do kladu Dinosauriformes, mierzącego około 60 cm długości, odkrytego i opisanego przez Friedricha von Huenego w 1910 roku. Żył w późnym triasie na terenie dzisiejszej Szkocji. Miał wydrążone i puste w środku kości, podobnie jak ptaki. Na podstawie porównań z innymi członkami Dinosauriformes wnioskuje się, że Saltopus miał pięciopalczaste dłonie oraz długą czaszkę z wieloma ostrymi zębami. Materiał kopalny jest jednak bardzo ubogi – obejmuje głównie kości kończyn.
 W
WSilezaur (Silesaurus) – rodzaj archozaura żyjącego w późnym triasie na terenie współczesnej Europy. Odkrycie skamieniałości co najmniej kilkunastu osobników pozwoliło na stosunkowo dokładną rekonstrukcję morfologii zwierzęcia, a także zbadanie mikrostruktury kości i fizjologii. Analizy te sugerują, że należał do grupy zwierząt zwinnych i aktywnych. Silezaur był prawdopodobnie średniej wielkości czworonożnym roślinożercą prowadzącym stadny tryb życia. Filogenetyczna pozycja rodzaju Silesaurus była przedmiotem intensywnych debat od chwili jego opisania. Niektórzy naukowcy klasyfikowali go jako prawdziwego dinozaura, jednak większość paleontologów uznaje go za bliską grupę zewnętrzną dinozaurów. Wszystkie znane skamieniałości należące bezsprzecznie do przedstawicieli tego rodzaju pochodzą z Krasiejowa w województwie opolskim.
 W
WSkleromochl (Scleromochlus) – rodzaj niewielkiego archozaura żyjącego w późnym triasie na obecnych terenach Europy. Dotychczas odnaleziono szczątki kilku osobników, co sprawia, że anatomia zwierzęcia nie jest w pełni poznana. Scleromochlus był prawdopodobnie dwunożnym biegaczem lub skoczkiem, zdolnym do skakania na duże odległości. Filogenetyczna pozycja rodzaju stanowiła przedmiot długich debat wśród paleontologów – zaliczano go zarówno do krokodylej linii archozaurów, jak również tej prowadzącej ku ptakom. Przedstawiono kilka teorii dotyczących jego sposobu poruszania się, jednak ze względu na niekompletność materiału kopalnego żadnej nie udało się w pełni potwierdzić. Wszystkie znane skamieniałości skleromochli pochodzą z formacji Lossiemouth Sandstone w Szkocji.
 W
WSmok – rodzaj dużego drapieżnego archozaura żyjącego w późnym triasie na terenach współczesnej Europy. Jego skamieniałości pochodzą z datowanych na przełom noryku i retyku osadów w cegielni w Lisowicach. Mierząc 5–6 m długości, z czego blisko 50–60 cm przypadało na czaszkę, był jednym z największych triasowych archozaurów. Jego pozycja filogenetyczna jest trudna do ustalenia – Smok wykazuje pewne cechy występujące u teropodów, rauizuchów i bardziej prymitywnych archozaurów.
 W
WSpondylosoma – znany z nielicznych skamieniałości archozaur o niepewnej pozycji systematycznej. Żył w środkowym lub późnym triasie na terenach dzisiejszej Ameryki Południowej. Jego szczątki znaleziono w Brazylii.
 W
WSzarowipteryks (Sharovipteryx) – rodzaj wczesnotriasowego gada, jednego z pierwszych gadów szybujących. Miał około 20 cm, z długim ogonem i ważył około 7,5 gramów. Mógł być krewnym lub przodkiem pterozaurów, chociaż ta teoria jest kontrowersyjna. Inaczej niż u pterozaurów, główna błona była rozpięta między tylnymi a nie przednimi nogami.
WTanystropheus, tanystrof – rodzaj gadów żyjących podczas środkowego triasu około 230 milionów lat temu. Miały 6 metrów długości, a wyróżniały się bardzo wydłużoną szyją, która mierzyła 3 metry, czyli tyle ile tułów i ogon razem wzięte. Pomimo tak długiej szyi miały tylko dziesięć kręgów szyjnych. Z powodu takiej budowy często przyjmuje się, że prowadziły wodny lub półwodny tryb życia, zamieszkując blisko brzegu i żywiąc się rybami lub skorupiakami. Jednak budowa ciała Tanystropheus nie wskazuje żeby miały jakiekolwiek cechy adaptacji do takiego trybu życia.
 W
WTasmaniosaurus triassicus jest nazwą gatunkową żyjącego we wczesnym triasie archozauromorfa z grupy Archosauriformes i rodziny Proterosuchidae, którego szczątki odkryto w formacji Knocklofty. Ponieważ szkielet zwierzęcia zachował się całkowicie, stanowi on nalepiej zachowane gadzie szczątki odnalezione na terenie Australii. Jest też jednym z najstarszych znanych gadów odnalezionych na terenie tego kontynentu. Inny przedstawiciel jego rodziny, kalizuch, żył we wczesnym triasie w okolicach dzisieszego Queensland.
 W
WTejuwazu – archozaur z kladu Dinosauriformes żyjący w późnym triasie (karnik) na terenach dzisiejszej Ameryki Południowej. Znany materiał kopalny to tylko holotyp – prawa kość udowa oznaczona BSPG AS XXV 53 o długości 27,6 cm i kość piszczelowa BSPG AS XXV 54 o długości 26,4 cm; skamieniałości te odkryto w osadach formacji Santa Maria. Teyuwasu znaczy "wielki jaszczur". Nie wiadomo nic pewnego na temat wielkości i wyglądu tego archozaura; zachowane kości sugerują, że był on większy i mocniej zbudowany od niektórych z najwcześniejszych znanych dinozaurów. Jego kość piszczelowa była wyraźnie dłuższa od znanych kości piszczelowych saturnalii i eoraptora i nieco dłuższa od kości piszczelowej staurikozaura, była natomiast krótsza od niektórych znanych kości piszczelowych herrerazaura.
 W
WTekodonty (Thecodontia) – parafiletyczna grupa wczesnych archozaurów o budowie pokrojowo przypominającej krokodyle, ale z bardziej prostymi nogami niż inne gady. Poza najprymitywniejszymi formami większość posiadała uzębienie tekodontyczne – zęby osadzone w zębodołach szczęki i żuchwy. Czaszka diapsydalna, zęby zwykle ostre i zakrzywione ku tyłowi.
 W
WTeraterpeton – rodzaj archozauromorfa żyjącego w późnym triasie na obecnych terenach Ameryki Północnej. Został opisany w 2003 roku przez Hansa-Dietera Suesa na podstawie niekompletnego szkieletu odkrytego w datowanych na karnik osadach formacji Wolfville w Nowej Szkocji. Obszar skroniowy czaszki teraterpetona jest euryapsydalny. Rostrum kości przedszczękowej oraz spojenie żuchwowe są wydłużone i bezzębne. Uzębienie składa się z odmiennych, nachodzących na siebie górnych i dolnych zębów. Według analizy filogenetycznej przeprowadzonej przez Suesa Teraterpeton należy do kladu Archosauromorpha i może być blisko spokrewniony z Trilophosaurus. Potwierdziły to również późniejsze analizy kladystyczne, według których Teraterpeton jest taksonem siostrzanym do pozostałych przedstawicieli Trilophosauridae, czyli Trilophosaurus i Spinosuchus. Teraterpeton to trzeci znany – po trilofozaurze i doswellii – późnotriasowy archozauromorf mający euryapsydalną czaszkę.
 W
WTrilophosaurus – roślinożerny gad z kladu Archosauromorpha, żyjącego w późnym triasie. Żył na obszarze równikowej Pangei, w jej zachodniej strefie.
WTrilofozaury (Trilophosauria) – rząd triasowych archozauromorfów. Obejmuje jedną rodzinę – Trilophosauridae. Były to średniej wielkości zwierzęta roślinożerne, przypominające nieco rynchozaury. Najlepiej poznany rodzaj – Trilophosaurus – osiągał do 3 m długości, z czego na czaszkę przypadało zaledwie 10–15 cm. Kość przedszczękowa i przednia część żuchwy były u trilofozaurów bezzębne i prawdopodobnie zakończone rogowym dziobem.
 W
WVancleavea – rodzaj archozauromorfa z kladu Archosauriformes żyjącego w późnym trasie na terenach dzisiejszej Ameryki Północnej. Holotypem jest niekompletny szkielet pozaczaszkowy oznaczony PEFO 2427, odkryty w osadach formacji Chinle na terenie Parku Narodowego Skamieniałego Lasu w Arizonie. Liczne skamieniałości Vancleavea odkryto w stanach Arizona, Nowy Meksyk, Teksas i Utah; prawie kompletny szkielet z zachowaną czaszką, oznaczony GR 138, odkryto na terenie Ghost Ranch w Nowym Meksyku. Wiek znanych skamieniałości wskazuje, że rodzaj Vancleavea przetrwał prawie 20 milionów lat, od karniku do retyku; gatunki zwykle nie istnieją tak długo, co może sugerować, że rodzaj Vancleavea obejmował więcej niż jeden gatunek. Najbardziej kompletne szkielety przedstawicieli tego rodzaju różnią się między sobą proporcjami niektórych kości, jednak obecnie nie da się stwierdzić, czy różnice te dowodzą, że szkielety należały do zwierząt z różnych gatunków czy zwierząt z tego samego gatunku w różnym wieku. Nesbitt i współpracownicy (2009) zaliczyli do rodzaju Vancleavea tylko jeden gatunek – gatunek typowy tego rodzaju, V. campi.
 W
WWenatikozuch (Venaticosuchus) – rodzaj archozaura zaliczanego do grupy Crurotarsi. Żył pod koniec triasu, około 220 milionów lat temu, na terenie obecnej Argentyny, której obszar wchodził wówczas w skład superkontynentu Pangei.
 W
WYounginiformes – grupa gadów z podrgromady diapsydów. Alfred Romer w 1947 roku zastąpił nazwę Eosuchia nazwą younginiformes.